Phase Transitions of Lipids and Liposomes
Introduction
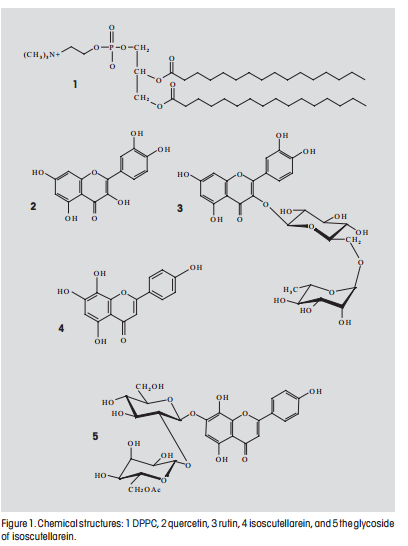
Phospholipid molecules consist of a polar head linked to two long acyl groups (e.g. DPPC, Fig. 1. 1). When dispersed in water, lipids align themselves with their polar heads toward the water to form micelles (clusters), liposomes (microscopic concentric spheres or vesicles) or other structures. Liposomes are produced when some phospholipids aggregate to form double layers of molecules and then close to form bilayer membranes.
Both unilamellar (SUVs, small unilamellar vesicles) and multilamellar (MLVs, multilamellar vesicles) structures can be produced into which drug molecules can be incorporated. This has led to the use of liposomes as drug delivery systems for medical applications (via intravenous injection). Liposomes resemble the membrane of a living cell and are used as models for cell membranes.
Flavonoids are polyphenolic compounds with a diphenylpropane structure (C6-C3-C6) that exhibit different biological effects, e.g. anti-inflammatory, bactericidal, antiviral and fungicidal action, tumor growth reduc-tion, protective effect for the liver, antitumor- and antioxidation activity [1]. In this study, several flavonoids have been incorporated into liposomes. These compounds are able to penetrate cell walls and their ability to alleviate cardiac diseases has been described in the literature [2]. The interaction of flavonoids with liposomes has been investigated to gain a better understanding of their interaction with cell membranes and to tailor-make liposomes as controlled-release drug delivery systems.
The DSC heating curves of lipid bilayers show two phase transitions: first a small peak, the so-called pretransition, followed by a second (main) transition.
DPPC bilayers exist in the gel phase (Lβ’ ) at temperatures below 35 °C, whereas above 42 °C they are present in the liquid crystalline phase (Lα’ ). The pretransition corresponds to a reorganization of individual lipid molecules in the lipid bilayer. Between 35 and 42 °C, the phospholipid bilayer is in the Pβ’ or so-called “rippled phase”. Following the pretransition at 35 °C, several conformational changes occur in the lipid molecules as well as changes in the geometry of the bilayers [5]. The most notable change is trans-gauche isomerization, which changes the acyl chain conformation (see Fig. 2). The number of gauche conformational isomers influences the fluidity. This depends not only on the temperature but also on the penetration of active substance molecules present, which changes the lipid environment. With a given polar head, the transition temperature depends on the type and length of the acyl chains – the longer the chain, the higher the temperature. Pure DPPC bilayers exhibit characteristic transitions with low enthalpy changes and a sharp main transition (Tm), both of which occur at the expected transition temperatures of 35.1 and 41.3 °C.
DSC is a useful technique to study the thermal effects of additives in bilayer membranes and has already been used in the past to study flavonoid-biomembrane interactions [3].
Materials and Methods
Liposome Preparation and Flavonoid Incorporation
Pure Liposomes
Liposomes were prepared using the thin film hydration method [4]. DPPC (66.06 mg) was weighed into a round-bottom flask and dissolved in chloroform. The solvent was distilled off at -40 °C leaving a thin film on the inner surface of the flask. The lipid film was hydrated with HPLC grade deionized water (3 mL) at 51 °C for one hour, which led to the formation of MLVs (multilamellar vesicles). A 300-µl portion of this material was used for the DSC measurements. The remainder of the sample was subjected to two cycles, each lasting 20 min with an interval of 5 min in a probe sonicator in an ice bath. This resulted in the formation of SUVs (small unilamellar vesicles). Some 500 µL of this was lyophilized and used for other DSC measurements. The MLVs and SUVs were prepared according to the method described above and afterward freeze-dried. Hydration of the stored powder regenerated the liposome suspensions.
Liposome-Flavonoid Mixtures
dissolved in ethanol. Rutin (Fig. 1. 3) (0.020 µmoles), isoscutellarein (Fig. 1. 4) (0.020 µmoles) and isoscutellarein glycoside (Fig. 1. 5) (0.020 µmoles) were dissolved in methanol. Each flavonoid solution was then added to a lipid solution and the organic solvent distilled off under reduced pressure. The samples were stored overnight in a desiccator. The lipid film was hydrated through the addition of HPLC grade water (3 mL) and the MLVs produced by vigorously shaking the suspensions in a water bath above Tm, the transition temperature from gel to the liquid crystalline phase of the lipid (41 °C), and stirring for 75 minutes. The resulting liposome suspensions consisting of MLVs were frozen in a dry ice bath (solid CO2/n-butanol) and thawed by heating to 41 °C in a water bath. The size of the vesicles was reduced by performing a series of 15 freezing/thawing cycles: The large unilamellar vesicles (LUVs) that resulted were subjected to ultrasound in an ice bath for two periods of 20 minutes with a pause of 5 minutes. The resulting vesicles were then held at room temperature for 30 minutes. Unbound flavonoids were removed at room temperature by gel filtration chromatography using a saturated Sephadex (G-75) column equilibrated with HPLC grade water (pH 5.6).
DSC Measurements
The MLV and SUV samples were measured using a METTLER TOLEDO DSC822e equipped with an HSS7 sensor and a Julabo intracooler. Pure MLV and SUV samples were prepared by dispersing the powdered liposomes in the appropriate quantities of bidistilled water.
Conclusions
DSC studies on the interaction of flavonoids with model DPPC membranes and on the relationship of the flavonoid structure to the lipid environment showed that the efficiency of incorporation depends on the structure of the flavonoid. The results indicate that the presence of a sugar group, or even just a number of hydroxyl groups in different positions in the flavonoid structure, plays a role in the incorporation of flavonoids in liposomes and the interaction of flavonoids with DPPC membranes.
The liposome formulations with flavonoids were also tested for their activity against three human cancer cell lines. In the case of quercetin, the formulation showed a lower degree of growth inhibition compared with the free quercetin; improved growth inhibition was however observed with the isoscutellarein formulation [5].
Phase Transitions of Lipids and Liposomes | Thermal Analysis Application No. UC 216 | Application published in METTLER TOLEDO Thermal Analysis UserCom 21